تکنولوژی زیستی :
تکنولوژی زیستی :
1-مهندسی ژنتیک :
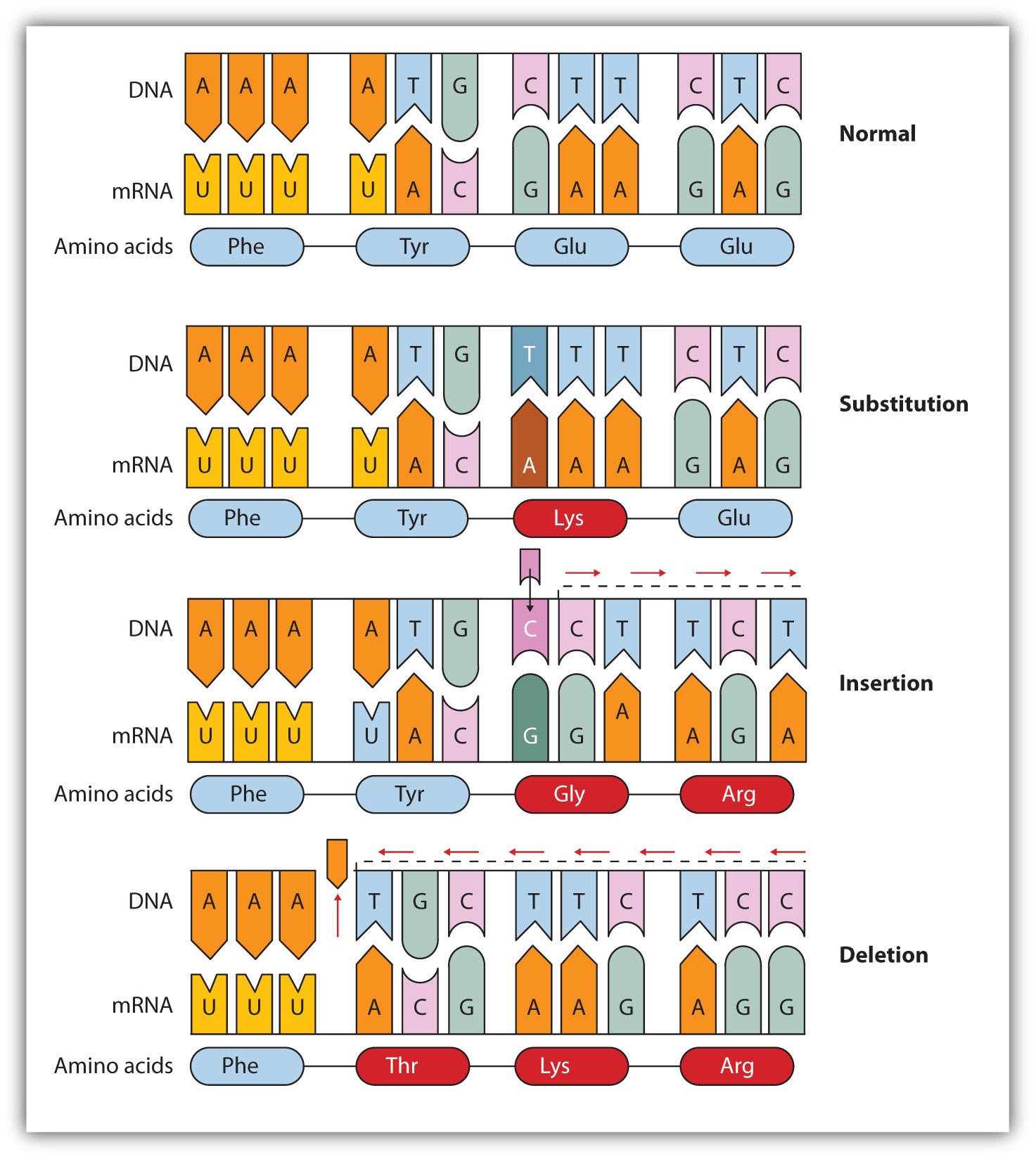
اولین بار در سال 1973 استانلی کوهن و هربرت بایر با طراحی و اجرا آزمایشی پژوهش های ژنتیک را متحول کرد. آنان ژن رمزکننده RNA ریبوزومی را از DNA قورباغه آفریقایی استخراج و به DNA باکتری اشریشیا کلای وارد کردند. در نتیجه باکتری هنگام رونویسی ، rRNA قورباغه را می سازد.
نکته : اولین جاندار دست ورزی شده باکتری اشریشیا کلای ( پروکاریوت ) می باشد.
مهندسی ژنتیک : استفاده از تکنیک های آزمایشگاهی برای ساخت مولکول DNA ی که حاوی ژن های جدید با ترکیب جدیدی از ژن هاست.
مهمترین هدف مهندسی ژنتیک : تولید ژن یا فرآورده آن به مقدار انبوه می باشد.برای اینکار بایستی ژن مورد نظر را از میان ژن های جاندار جدا و سپس آنرا به جاندار ساده ای مثل باکتری که تولید مثل سریعی دارد وارد کرده تا ژنهای مورد نظر توسط باکتری همانندسازی و به مقدار زیاد تهیه گردد.
ابزارهای مورد نیاز عبارتند از :
الف ) ابزاری که DNA را ببرد ( آنزیم های محدود کننده )
ب ) وسیله ای برای حمل ژن مورد نظر به درون باکتری
ج ) ابزاری برای ایجاد پیوند بین ژن خارجی و DNA باکتری
روش ها و ابزارهای مهندسی ژنتیک :
وکتور : عاملی برای انتقال ژن مرد نظر به سلول های میزبان در مهندسی ژنتیک
برخی از انواع وکتور عبارتند از : پلازمیدها و ویروس ها ( باکتریوفاژ )
پلازمید : مولکول DNAحلقوی کوچک که در برخی باکتریها وجود دارد و حاوی ژن های است که در کروموزوم اصلی باکتری وجود ندارد مانند ژن مقاومت به آنتی بیوتیک ( کروموزوم کمکی ) ، همانند سازی آن مستقل از کروموزوم اصلی بوده و حتی زمانی که باکتری در حال تولید مثل نیست نیز می تواند همانند سازی کند.
نکته : مهندسان ژنتیک ، ژن مورد نظر را درون پلازمید قرار می دهند.بنابراین هرگاه پلازمید همانند سازی نماید ژن مورد نظر را نیز همانند سازی خواهد نمود .
ژن خارجی : ژنی که از DNA جانداری استخراج و به DNA جانداری دیگر مانند باکتری اضافه می گردد.
DNA نوترکیب : DNA ای که از ترکیب دو DNA متفاوت ، ژن خارجی و دیگری پلازمید ( وکتور ) حاصل شده است .
نکته : از آنجائیکه یکی از اصلی ترین مراحل مهندسی ژنتیک ، تولید DNA نوترکیب می باشد ، مهندسی ژنتیک را فناوری DNA نوترکیب نیز می نامند.
باکتریوفاژ : ویروس هایی که میزبان آنها باکتری می باشد. با قرار دادن ژن خارجی در DNA باکتریوفاژ ، امکان تکثیر ژن فراهم می شود.
ساختن مولکول DNA نوترکیب : برای اینکار به دو آنزیم زیر نیاز داریم ک
الف ) آنزیمی برای بریدن پلازمید یا وکتور ( آنزیم محدودکننده )
ب ) آنزیمی برای اتصال دو سر ژن خارجی به پلازمید ( آنزیم لیگاز )
نکته : منظور از بریدن DNA قطع پیوند فسفودی استر و منظور از اتصال دو DNA برقراری پیوند فسفو دی استر میان دو DNA است .
آنزیم های محدودکننده : آنزیم هایی باکتریایی که توالی کوتاه و خاصی از DNA ( جایگاه تشخیص آنزیم ) را شناسایی کرده و سپس آنرا برش می دهند. مانند آنزیم EcoRI که توالی زیر را شناسایی می کنند.

جایگاه تشخیص آنزیم : محدود کننده توالی خاص از DNA که آنزیم آنرا شناسایی می کند.
نکته : توالی دو رشته جایگاه تشخیص عکس یکدیگر هستند.
بیشتر آنزیم های محدود کننده ، قطعاتی از DNA کوتاه تک رشته ای در هر دو انتها تولید می کنند که با یکدیگر مکمل هستند . این دو انتها ، انتهای چسبنده نامیده می شود. برخی نیز انتهای غیر چسبنده تولید می کنند که برای ما کارآیی ندارد.

نکته : وکتورهای به کار برده شده ، فقط دارای یک جایگاه تشخیص آنزیم هستند.
نکته : آنزیم محدود کننده ای که برای بریدن پلازمید استفاده می شود ، باید همان آنزیمی باشد که دو سر ژن خارجی با آن بریده شده است.در این صورت ، انتهای چسبنده یکی به به انتهای چسبنده دیگری متصل می شود.( اتصال توسط پیوندهای هیدروژنی است )
نکته : برای برقراری پیوند فسفو دی استر میان دو DNA ، مهندسان ژنتیک از آنزیمی به نام لیگاز استفاده می شود.
کلون شدن ژن : هنگامیکه DNA نوترکیب وارد باکتری شود ، باکتری با استفاده از دستگاه همانند سازی خود نسخه های متعددی از آن و در نتیجه ژن خارجی می سازد . ایجاد نسخه های متعدد و یکسان از ژن را کلون کردن ژن گویند.
نکته :همه باکتریها موفق به جذب DNA نوترکیب نمی شوند.
غربال کردن : در این مرحله باکتریهایی که DNA نوترکیب را جذب کرده ، از باکتریهایی که آنها را جذب نکرده اند جدا می کنند.
نکته : برای این کار از آنتی بیوتیک ها استفاده می شود. باکتریهای که DNA نوترکیب را جذب نکرده اند توسط آنتی بیوتیک از بین می روند. ژن مقاومت به آنتی یوتیک در پلازمید وجود دارد.
استخراج ژن : در این مرحله ژن خارجی از DNA نوترکیب جدا می شود. برای اینکار از همان آنزیم محدوکننده ای استفاده می شود که برای تولید DNA نوترکیب استفاده شده است . چون دقیقا از نقاطی که به هم وصل گردیده جدا شوند.
برای تفکیک پلازمید و ژن خارجی از دستگاه الکتروفورز استفاده می شود.که در آن ذرات بر اساس بار الکتریکی و اندازه در طول ژل از یکدیگر جدا می گردند.
نکته : DNA بار منفی دارد و به سمت قطب مثبت حرکت می کند.
نکته : مولکول های کوچکتر ( ژن خارجی ) سریعتر از مولکول های بزرگ ( پلازمید ) از منافذ عبور می کنند و به قطب مثبت نزدیکترند.
نکته : روش الکتروفورز علاوه بر نوکلئیک اسیدها ، برای پروتئین ها نیز کاربرد دارد. در این روش ، پروتئین ها بر اساس اندازه ، از یکدیگر جدا می شوند.
مهندسی ژنتیک در پزشکی :
داروها و واکسن های حاصل از مهندسی ژنتیک امروزه در دسترس هستند.
داروها : بسیاری از بیماریهای ژنی به علت عدم توانایی بدن در ساختن یک نوع پروتئین خاص است که با تولید آن توسط مهندسی ژنتیک می توان بیماریهای مزبور را درمان نمود. مانند مواد ضد انعقاد خون یا انسولین
فاکتور انعقادی شماره هشت ، پروتئینی است که در روند انعقاد خون نقش ددارد و فقدان آن باعث بیماری هموفیلی می گردد. امروزه این فاکتور به روش مهندسی ژنتیک ساخته می شود.
نکته : در گذشته فاکتور انعقادی از خون های اهدایی استخراج می گردید و انتقال آن به افراد نیازمند گاها باعث ابتلاء به ویروس HIV یا هپاتیت می گردید. اولین مورد ایدزشناخته شده در ایران از این طریق بوده است.
واکسن ها : بسیاری از بیماریها ی ویروسی ، مانند آبله و فلج اطفال با داروهای موجود درمان نمی شوند. بنابراین با به کار بردن واکسنه ها می توان از آنها پیشگیری نمود.
واکسن : میکروب کشته شده یا ضعیف شده یا سم خنثی شده میکروب و یا آنتی ژن های سطحی باکتری که به بدن تزریق و باعث بروز پاسخ ایمنی می گردد.
نکته : مشکل واکسن های قدیمی اینست که یک خطا در کشتن یا ضعیف کردن یک عامل بیماریزا منجر به انتقال بیماری به افرادی می شود که برای جلوگیری از آن اقدام کرده اند.
واکسنهایی که با روش مهندسی ژنتیک ساخته می شوند این خطر را ندارند. در این روش ژن مربوط به آنتی ژن های یک عامل بیماریزا را به DNA یک باکتری یا ویروس غیر بیماریزا وارد می کنند .تا ویروس یا بکتری غیر بیماریزا آنتی ژنهایی سطحی عامل بیماریزا را تولید کنند و باعث بروز پاسخ ایمنی شوند.
نکته : عامل ایجاد بیماری آنتی ژن هایی سطحی نیست بلکه فقط به کمک آنها توسط دستگاه ایمنی تشخیص داده می شوند.
مانند تولید واکسن هرپس تناسلی یا واکسن هپاتیت B ، امروزه تلاش گردیده تا در مقابل مالاریا واکسنی موثر تولید گردد.
ژن درمانی : قرار دادن یک نسخه سالم از یک ژن ، درون سلول های فردی که دارای نسخه ناقص از آن ژن است.
نکته : بسیاری از ناهنجاریهای ژنتیک بعلت عدم وجود نسخه فعال یک ژن خاص در بدن می باشد.
در این روش سلول ها را از بدن بیمار خارج و ژن سالم را وارد آنها می کنند. سپس سلول های تغییر یافته را به بدن بیمار باز می گردانند. پس از آن ماده ای که در این فرد وجود نداشت ، توسط سلول های دارای ژن جدید ، ساخته می شود.
در حال حاضر ژن درمانی در سلول هایی قابل استفاده است که قدرت تقسیم آن سلول ها و یا سلول های مادر( پایه ای ) آنها بسیار زیاد باشد. مثلا در مورد سلو لهای خونی و پروتئین های حاصل از آنها ، چون منشاء آنها سلولهای مغز استخوان است که قدرت تقسیم میتوز بالا دارند.
نکته : اولین تلاش ها برای ژن درمانی در مورد دختر بچه ای مبتلائ به نوعی ناهنجاری دستگاه ایمنی صورت گرفت.علت بیماری ژن جهش یافته ای بود که نمی توانست یک آنزیم مهم دستگاه ایمنی را بسازد.
توالی و جایگاه همه ژن های انسان مورد مطالعه قرار گرفته است :

پروژه ژنوم انسان HGP : هدف از آن تعیین توالی نوکلئوتیدهای ژنوم انسان و تعیین نقشه جایگاه هر ژن روی هر کروموزوم است.
ژنوم : کل محتوای DNA یک جاندار که شامل محتوای DNA هسته ای و DNA های سیتوپلاسمی ( میتوکندری و کلروپلاست ) می باشد.
نکته : دانشمندان امیدوارند با دانش به دست آمده از پروژه ژنوم انسان بتوانند ، حدود 4000 بیماری ژنتیک انسان را تشخیص معالجه و درمان نمایند.
نکته : دانشمندان تاکنون ژنهای بیماری های زیادی از جمله سیستیک فیبروزیز ، دیستروفی عضلانی دوشن و سرطان را کشف نموده اند. بیش از 450 ژن و 200 ناهنجاری کروموزومی روی کروموزوم X وجود دارد ( وابسته به جنس ) مانند موارد زیر :
تحلیل عضلانی دوشن ، رنگدانه ای شدن شبکیه چشم ، سیناپسین 1 ، کام شکافدار وابسته به X ، پذیرنده آنژیوتانسین 2 ، نشانگان زالی – ناشنوایی و پروتئین ریبوزومی 10 L
مهندسی ژنتیک در کشاورزی و دامداری :
می توان با انتقال ژن ها به گیاهان باعث اصلاح محصولات آنها شد.
امروزه مهندسان ژنتیک می توانند ویژگی های مطلوب را با دست ورزی ژن به گیاهان بیفزایند. مثلا ایجاد گیاهان مقاوم به شرایط خشکی و تولید گیاهانی که با خاک های مختلف ، اقلیم های متفاوت و فشارهای محیطی سازگاری حاصل کنند.تنظیم سرعت رسیدن میوه ها و افزایش ارزش غذایی گیاهان.
نکته : با انجام روش های مهندسی ژنتیک روی گیاه برنج ، سویه های دارای میزان بالای کاروتن ( که در بدن به ویتامین A تبدیل می شود ) و آهن تولید شده اند . این امر برای مردم بخش های از قاره آسیا که کمبود این دو ماده را دارند بسیار مهم است.
دانشمندان با وارد کردن یک ژن درون محصولات گیاهی ، گیاهانی تولید کرده اند که نسبت به حشرات مقاوم هستند. گیاهانی که نسبت به حشرات مقاوم اند، نیازی به استفاده از سموم حشره کش آلوده کننده محیط زیست ندارند.
کشف یک وکتور گیاهی :

تا چندین سال ، مهندسان ژنتیک وکتور مناسبی که بتواند ژن ها را به گیاه انتقال دهد، در دسترس نداشتند
پلازمید Ti : نوعی پلازمید باکتریایی که باعث ایجاد تومور در سلول های گیاهی می شود ( القاء کننده ایجاد تومور )، پلازمید Ti بسیاری از گیاهان زراعی مثل گوجه فرنگی ، توتون و سویا را آلوده می کند.
گال : نوعی ببیماری گیاهی که باعث ایجاد تومورهای بزرگ روی گیاه می شود.
نکته : محققان ژن ایجاد کننده تومور را از پلازمید Ti خارج و یک DNA خاص ( ژن خارجی ) را جایگزین آن می کنند.
نکته : می توان ژن را با یک تفنگ ژنی به سلول های گیاه گندم شلیک کرد.
تکنولوژی ژن در دامداری به کار برده می شود.در گذشته دامهایی که شیر یا سایر محصولات فراوانی دارند را انتخاب و پرورش می دانند.این فرآیند ها ی متوالی ، طولانی و کم بازده است. امروزه از روش های مهندسی ژنتیک برای اصلاح یا تغییر دام ها استفاده می شود.
نکته : برخی از دامداران برای افزایش تولید شیر به رژیم غذایی گاوها هورمون های رشد که از مغز گاوهای کشته شده استخراج می شد ، افزوده می شد ، امروزه هورمون رشد گاوی به روش مهندسی ژنتیک توسط باکتری ها ساخته می شود.که هزینه آن کمتر و به صزفه تر است.
تولید پروتئین های مفید از نظر پزشکی : کاربرد دیگر تکنولوژی ژن در دامداری افزودن ژن های انسان به دام ها ست.هدف از این کار آن است که پروتئین های اانسان در شیر دام ها ظاهر شود. این روش بیشتر برای پرروتئین های پیچیده انسانی به کار می روندکه از طریق تکنولوژی ژن در باکتری ها تولید نمی شوند.
جانوران تراژن : جانورانی که در سلول های آنها، DNA بیگانه وجود دارد.
کلون کردن از سلو لهای تخصص یافته : اولین بار توسط یان ویلموت در سال 1997 ارائه گردید.که در آن یک بره ،با کلون کردن هسته سلولی از پستان گوسفند بالغ به وجود آمد.
نکته : در گذشته محققان تصور می کردند نمی توان از سلول های تمایز یافته برای تولید موجود زنده کامل استفاده کنند.
ویلموت سلول پستان گوسفند را در اثر تحریک الکتریکی با سلول تخمک فاقد هسته یک گوسفند دیگر ادغام کرد . این سلول ادغام شده تقسیم شد و اولین سلول های جنین را بوجود آورد. ویلموت سلول های حاصل را درون رحم گوسفند ماده ای ( مادر جانشینی ) کار گذاشت. حاصل این کار تولد بره دالی بود که از نظر ژنتیکی مشابه گوسفندی بود که سلول پستان از آن گرفته شده بود.
این کار توسط محققان دیگر برای کلون کردن گاوها و موش ها به کار رفته است. این آزمایش ها نشان می دهند که کلون کردن بدین روش در سایر پستانداران امکان پذیر است.
1-مهندسی ژنتیک :
اولین بار در سال 1973 استانلی کوهن و هربرت بایر با طراحی و اجرا آزمایشی پژوهش های ژنتیک را متحول کرد. آنان ژن رمزکننده RNA ریبوزومی را از DNA قورباغه آفریقایی استخراج و به DNA باکتری اشریشیا کلای وارد کردند. در نتیجه باکتری هنگام رونویسی ، rRNA قورباغه را می سازد.
نکته : اولین جاندار دست ورزی شده باکتری اشریشیا کلای ( پروکاریوت ) می باشد.
مهندسی ژنتیک : استفاده از تکنیک های آزمایشگاهی برای ساخت مولکول DNA ی که حاوی ژن های جدید با ترکیب جدیدی از ژن هاست.
مهمترین هدف مهندسی ژنتیک : تولید ژن یا فرآورده آن به مقدار انبوه می باشد.برای اینکار بایستی ژن مورد نظر را از میان ژن های جاندار جدا و سپس آنرا به جاندار ساده ای مثل باکتری که تولید مثل سریعی دارد وارد کرده تا ژنهای مورد نظر توسط باکتری همانندسازی و به مقدار زیاد تهیه گردد.
ابزارهای مورد نیاز عبارتند از :
الف ) ابزاری که DNA را ببرد ( آنزیم های محدود کننده )
ب ) وسیله ای برای حمل ژن مورد نظر به درون باکتری
ج ) ابزاری برای ایجاد پیوند بین ژن خارجی و DNA باکتری
روش ها و ابزارهای مهندسی ژنتیک :
وکتور : عاملی برای انتقال ژن مرد نظر به سلول های میزبان در مهندسی ژنتیک
برخی از انواع وکتور عبارتند از : پلازمیدها و ویروس ها ( باکتریوفاژ )
پلازمید : مولکول DNAحلقوی کوچک که در برخی باکتریها وجود دارد و حاوی ژن های است که در کروموزوم اصلی باکتری وجود ندارد مانند ژن مقاومت به آنتی بیوتیک ( کروموزوم کمکی ) ، همانند سازی آن مستقل از کروموزوم اصلی بوده و حتی زمانی که باکتری در حال تولید مثل نیست نیز می تواند همانند سازی کند.
نکته : مهندسان ژنتیک ، ژن مورد نظر را درون پلازمید قرار می دهند.بنابراین هرگاه پلازمید همانند سازی نماید ژن مورد نظر را نیز همانند سازی خواهد نمود .
ژن خارجی : ژنی که از DNA جانداری استخراج و به DNA جانداری دیگر مانند باکتری اضافه می گردد.
DNA نوترکیب : DNA ای که از ترکیب دو DNA متفاوت ، ژن خارجی و دیگری پلازمید ( وکتور ) حاصل شده است .
نکته : از آنجائیکه یکی از اصلی ترین مراحل مهندسی ژنتیک ، تولید DNA نوترکیب می باشد ، مهندسی ژنتیک را فناوری DNA نوترکیب نیز می نامند.
باکتریوفاژ : ویروس هایی که میزبان آنها باکتری می باشد. با قرار دادن ژن خارجی در DNA باکتریوفاژ ، امکان تکثیر ژن فراهم می شود.
ساختن مولکول DNA نوترکیب : برای اینکار به دو آنزیم زیر نیاز داریم ک
الف ) آنزیمی برای بریدن پلازمید یا وکتور ( آنزیم محدودکننده )
ب ) آنزیمی برای اتصال دو سر ژن خارجی به پلازمید ( آنزیم لیگاز )
نکته : منظور از بریدن DNA قطع پیوند فسفودی استر و منظور از اتصال دو DNA برقراری پیوند فسفو دی استر میان دو DNA است .
آنزیم های محدودکننده : آنزیم هایی باکتریایی که توالی کوتاه و خاصی از DNA ( جایگاه تشخیص آنزیم ) را شناسایی کرده و سپس آنرا برش می دهند. مانند آنزیم EcoRI که توالی زیر را شناسایی می کنند.
جایگاه تشخیص آنزیم : محدود کننده توالی خاص از DNA که آنزیم آنرا شناسایی می کند.
نکته : توالی دو رشته جایگاه تشخیص عکس یکدیگر هستند.
بیشتر آنزیم های محدود کننده ، قطعاتی از DNA کوتاه تک رشته ای در هر دو انتها تولید می کنند که با یکدیگر مکمل هستند . این دو انتها ، انتهای چسبنده نامیده می شود. برخی نیز انتهای غیر چسبنده تولید می کنند که برای ما کارآیی ندارد.
نکته : وکتورهای به کار برده شده ، فقط دارای یک جایگاه تشخیص آنزیم هستند.
نکته : آنزیم محدود کننده ای که برای بریدن پلازمید استفاده می شود ، باید همان آنزیمی باشد که دو سر ژن خارجی با آن بریده شده است.در این صورت ، انتهای چسبنده یکی به به انتهای چسبنده دیگری متصل می شود.( اتصال توسط پیوندهای هیدروژنی است )
نکته : برای برقراری پیوند فسفو دی استر میان دو DNA ، مهندسان ژنتیک از آنزیمی به نام لیگاز استفاده می شود.
کلون شدن ژن : هنگامیکه DNA نوترکیب وارد باکتری شود ، باکتری با استفاده از دستگاه همانند سازی خود نسخه های متعددی از آن و در نتیجه ژن خارجی می سازد . ایجاد نسخه های متعدد و یکسان از ژن را کلون کردن ژن گویند.
نکته :همه باکتریها موفق به جذب DNA نوترکیب نمی شوند.
غربال کردن : در این مرحله باکتریهایی که DNA نوترکیب را جذب کرده ، از باکتریهایی که آنها را جذب نکرده اند جدا می کنند.
نکته : برای این کار از آنتی بیوتیک ها استفاده می شود. باکتریهای که DNA نوترکیب را جذب نکرده اند توسط آنتی بیوتیک از بین می روند. ژن مقاومت به آنتی یوتیک در پلازمید وجود دارد.
استخراج ژن : در این مرحله ژن خارجی از DNA نوترکیب جدا می شود. برای اینکار از همان آنزیم محدوکننده ای استفاده می شود که برای تولید DNA نوترکیب استفاده شده است . چون دقیقا از نقاطی که به هم وصل گردیده جدا شوند.
برای تفکیک پلازمید و ژن خارجی از دستگاه الکتروفورز استفاده می شود.که در آن ذرات بر اساس بار الکتریکی و اندازه در طول ژل از یکدیگر جدا می گردند.
نکته : DNA بار منفی دارد و به سمت قطب مثبت حرکت می کند.
نکته : مولکول های کوچکتر ( ژن خارجی ) سریعتر از مولکول های بزرگ ( پلازمید ) از منافذ عبور می کنند و به قطب مثبت نزدیکترند.
نکته : روش الکتروفورز علاوه بر نوکلئیک اسیدها ، برای پروتئین ها نیز کاربرد دارد. در این روش ، پروتئین ها بر اساس اندازه ، از یکدیگر جدا می شوند.
مهندسی ژنتیک در پزشکی :
داروها و واکسن های حاصل از مهندسی ژنتیک امروزه در دسترس هستند.
داروها : بسیاری از بیماریهای ژنی به علت عدم توانایی بدن در ساختن یک نوع پروتئین خاص است که با تولید آن توسط مهندسی ژنتیک می توان بیماریهای مزبور را درمان نمود. مانند مواد ضد انعقاد خون یا انسولین
فاکتور انعقادی شماره هشت ، پروتئینی است که در روند انعقاد خون نقش ددارد و فقدان آن باعث بیماری هموفیلی می گردد. امروزه این فاکتور به روش مهندسی ژنتیک ساخته می شود.
نکته : در گذشته فاکتور انعقادی از خون های اهدایی استخراج می گردید و انتقال آن به افراد نیازمند گاها باعث ابتلاء به ویروس HIV یا هپاتیت می گردید. اولین مورد ایدزشناخته شده در ایران از این طریق بوده است.
واکسن ها : بسیاری از بیماریها ی ویروسی ، مانند آبله و فلج اطفال با داروهای موجود درمان نمی شوند. بنابراین با به کار بردن واکسنه ها می توان از آنها پیشگیری نمود.
واکسن : میکروب کشته شده یا ضعیف شده یا سم خنثی شده میکروب و یا آنتی ژن های سطحی باکتری که به بدن تزریق و باعث بروز پاسخ ایمنی می گردد.
نکته : مشکل واکسن های قدیمی اینست که یک خطا در کشتن یا ضعیف کردن یک عامل بیماریزا منجر به انتقال بیماری به افرادی می شود که برای جلوگیری از آن اقدام کرده اند.
واکسنهایی که با روش مهندسی ژنتیک ساخته می شوند این خطر را ندارند. در این روش ژن مربوط به آنتی ژن های یک عامل بیماریزا را به DNA یک باکتری یا ویروس غیر بیماریزا وارد می کنند .تا ویروس یا بکتری غیر بیماریزا آنتی ژنهایی سطحی عامل بیماریزا را تولید کنند و باعث بروز پاسخ ایمنی شوند.
نکته : عامل ایجاد بیماری آنتی ژن هایی سطحی نیست بلکه فقط به کمک آنها توسط دستگاه ایمنی تشخیص داده می شوند.
مانند تولید واکسن هرپس تناسلی یا واکسن هپاتیت B ، امروزه تلاش گردیده تا در مقابل مالاریا واکسنی موثر تولید گردد.
ژن درمانی : قرار دادن یک نسخه سالم از یک ژن ، درون سلول های فردی که دارای نسخه ناقص از آن ژن است.
نکته : بسیاری از ناهنجاریهای ژنتیک بعلت عدم وجود نسخه فعال یک ژن خاص در بدن می باشد.
در این روش سلول ها را از بدن بیمار خارج و ژن سالم را وارد آنها می کنند. سپس سلول های تغییر یافته را به بدن بیمار باز می گردانند. پس از آن ماده ای که در این فرد وجود نداشت ، توسط سلول های دارای ژن جدید ، ساخته می شود.
در حال حاضر ژن درمانی در سلول هایی قابل استفاده است که قدرت تقسیم آن سلول ها و یا سلول های مادر( پایه ای ) آنها بسیار زیاد باشد. مثلا در مورد سلو لهای خونی و پروتئین های حاصل از آنها ، چون منشاء آنها سلولهای مغز استخوان است که قدرت تقسیم میتوز بالا دارند.
نکته : اولین تلاش ها برای ژن درمانی در مورد دختر بچه ای مبتلائ به نوعی ناهنجاری دستگاه ایمنی صورت گرفت.علت بیماری ژن جهش یافته ای بود که نمی توانست یک آنزیم مهم دستگاه ایمنی را بسازد.
توالی و جایگاه همه ژن های انسان مورد مطالعه قرار گرفته است :
پروژه ژنوم انسان HGP : هدف از آن تعیین توالی نوکلئوتیدهای ژنوم انسان و تعیین نقشه جایگاه هر ژن روی هر کروموزوم است.
ژنوم : کل محتوای DNA یک جاندار که شامل محتوای DNA هسته ای و DNA های سیتوپلاسمی ( میتوکندری و کلروپلاست ) می باشد.
نکته : دانشمندان امیدوارند با دانش به دست آمده از پروژه ژنوم انسان بتوانند ، حدود 4000 بیماری ژنتیک انسان را تشخیص معالجه و درمان نمایند.
نکته : دانشمندان تاکنون ژنهای بیماری های زیادی از جمله سیستیک فیبروزیز ، دیستروفی عضلانی دوشن و سرطان را کشف نموده اند. بیش از 450 ژن و 200 ناهنجاری کروموزومی روی کروموزوم X وجود دارد ( وابسته به جنس ) مانند موارد زیر :
تحلیل عضلانی دوشن ، رنگدانه ای شدن شبکیه چشم ، سیناپسین 1 ، کام شکافدار وابسته به X ، پذیرنده آنژیوتانسین 2 ، نشانگان زالی – ناشنوایی و پروتئین ریبوزومی 10 L
مهندسی ژنتیک در کشاورزی و دامداری :
می توان با انتقال ژن ها به گیاهان باعث اصلاح محصولات آنها شد.
امروزه مهندسان ژنتیک می توانند ویژگی های مطلوب را با دست ورزی ژن به گیاهان بیفزایند. مثلا ایجاد گیاهان مقاوم به شرایط خشکی و تولید گیاهانی که با خاک های مختلف ، اقلیم های متفاوت و فشارهای محیطی سازگاری حاصل کنند.تنظیم سرعت رسیدن میوه ها و افزایش ارزش غذایی گیاهان.
نکته : با انجام روش های مهندسی ژنتیک روی گیاه برنج ، سویه های دارای میزان بالای کاروتن ( که در بدن به ویتامین A تبدیل می شود ) و آهن تولید شده اند . این امر برای مردم بخش های از قاره آسیا که کمبود این دو ماده را دارند بسیار مهم است.
دانشمندان با وارد کردن یک ژن درون محصولات گیاهی ، گیاهانی تولید کرده اند که نسبت به حشرات مقاوم هستند. گیاهانی که نسبت به حشرات مقاوم اند، نیازی به استفاده از سموم حشره کش آلوده کننده محیط زیست ندارند.
کشف یک وکتور گیاهی :
تا چندین سال ، مهندسان ژنتیک وکتور مناسبی که بتواند ژن ها را به گیاه انتقال دهد، در دسترس نداشتند
پلازمید Ti : نوعی پلازمید باکتریایی که باعث ایجاد تومور در سلول های گیاهی می شود ( القاء کننده ایجاد تومور )، پلازمید Ti بسیاری از گیاهان زراعی مثل گوجه فرنگی ، توتون و سویا را آلوده می کند.
گال : نوعی ببیماری گیاهی که باعث ایجاد تومورهای بزرگ روی گیاه می شود.
نکته : محققان ژن ایجاد کننده تومور را از پلازمید Ti خارج و یک DNA خاص ( ژن خارجی ) را جایگزین آن می کنند.
نکته : می توان ژن را با یک تفنگ ژنی به سلول های گیاه گندم شلیک کرد.
تکنولوژی ژن در دامداری به کار برده می شود.در گذشته دامهایی که شیر یا سایر محصولات فراوانی دارند را انتخاب و پرورش می دانند.این فرآیند ها ی متوالی ، طولانی و کم بازده است. امروزه از روش های مهندسی ژنتیک برای اصلاح یا تغییر دام ها استفاده می شود.
نکته : برخی از دامداران برای افزایش تولید شیر به رژیم غذایی گاوها هورمون های رشد که از مغز گاوهای کشته شده استخراج می شد ، افزوده می شد ، امروزه هورمون رشد گاوی به روش مهندسی ژنتیک توسط باکتری ها ساخته می شود.که هزینه آن کمتر و به صزفه تر است.
تولید پروتئین های مفید از نظر پزشکی : کاربرد دیگر تکنولوژی ژن در دامداری افزودن ژن های انسان به دام ها ست.هدف از این کار آن است که پروتئین های اانسان در شیر دام ها ظاهر شود. این روش بیشتر برای پرروتئین های پیچیده انسانی به کار می روندکه از طریق تکنولوژی ژن در باکتری ها تولید نمی شوند.
جانوران تراژن : جانورانی که در سلول های آنها، DNA بیگانه وجود دارد.
کلون کردن از سلو لهای تخصص یافته : اولین بار توسط یان ویلموت در سال 1997 ارائه گردید.که در آن یک بره ،با کلون کردن هسته سلولی از پستان گوسفند بالغ به وجود آمد.
نکته : در گذشته محققان تصور می کردند نمی توان از سلول های تمایز یافته برای تولید موجود زنده کامل استفاده کنند.
ویلموت سلول پستان گوسفند را در اثر تحریک الکتریکی با سلول تخمک فاقد هسته یک گوسفند دیگر ادغام کرد . این سلول ادغام شده تقسیم شد و اولین سلول های جنین را بوجود آورد. ویلموت سلول های حاصل را درون رحم گوسفند ماده ای ( مادر جانشینی ) کار گذاشت. حاصل این کار تولد بره دالی بود که از نظر ژنتیکی مشابه گوسفندی بود که سلول پستان از آن گرفته شده بود.
این کار توسط محققان دیگر برای کلون کردن گاوها و موش ها به کار رفته است. این آزمایش ها نشان می دهند که کلون کردن بدین روش در سایر پستانداران امکان پذیر است.
+ نوشته شده در دوشنبه بیستم آذر ۱۳۹۱ ساعت 23:14 توسط محمد جباری
|




























 محمد جباری
محمد جباری